
减数分裂是真核生物产生单倍体配子和进行有性生殖的重要生命过程。重组是减数分裂的核心事件,其过程高度复杂且受到严密调控。重组产生的交换不仅增加了后代的遗传多样性,同时也是作物遗传育种的重要基础。增加交换有助于缩短作物育种周期,而减少甚至消除交换有望应用于杂种优势的固定。因此,解析重组交换形成的分子调控机制具有重要的理论意义和应用价值。
RAD51旁系同源蛋白包括五个成员,分别为RAD51B、RAD51C、RAD51D、XRCC3和XRCC2。在哺乳动物中这些蛋白的编码基因突变往往引发胚胎致死,而在植物中这些基因突变影响植株育性却不影响营养生长,这就为研究RAD51旁系同源蛋白在减数分裂中的功能提供了重要材料。本文作者的前期研究证明RAD51旁系同源蛋白在重组早期扮演重要角色,负责将重组酶RAD51和DMC1招募到染色体上,并促进同源染色体配对和DNA单链入侵 (Zhang et al., 2020)。然而,RAD51旁系同源蛋白在重组晚期是否发挥功能依然未知。
2023年7月11日,565net必赢张凡凡博士在植物学Top期刊New Phytologist在线发表了题为“RAD51C-RAD51D interplays with MSH5 and regulates crossover maturation in rice meiosis”的文章,揭示了RAD51旁系同源蛋白在减数分裂中一个尚未发现的功能,即RAD51C-RAD51D复合体在重组晚期与交换形成的关键蛋白MSH5协同作用调控交换的成熟过程。
该研究利用CRISPR/cas9基因编辑技术对水稻RAD51C和RAD51D基因进行编辑并筛选到三个新的突变体:rad51c-3、rad51d-4和rad51d-5。不同于已发表的强等位突变体,这三个新突变体中期I和后期I染色体几乎不存在缠绕和碎片,而是呈二价体和单价体共存的表型,暗示这些突变体中交换形成受限 (图 1)。交叉结频率和交换指示蛋白HEI10的数目显著减少,表明RAD51C和RAD51D参与交换的成熟过程。RAD51D与交换形成相关蛋白MSH5之间的相互作用进一步说明RAD51旁系同源蛋白与MSH5协同调控交换前体的稳定和解开。此外,在rad51c-3、rad51d-4、rad51d-5、rad51c-3 rad51d-4、rad51c-3 dmc1a dmc1b和rad51d-4 dmc1a dmc1b这些单、双和三突变体中,染色体的缠绕程度随重组酶RAD51信号数量的减少而增加,说明在染色体上保留一定水平的RAD51蛋白是RAD51旁系同源蛋白发挥参与交换形成的晚期功能所必需的。
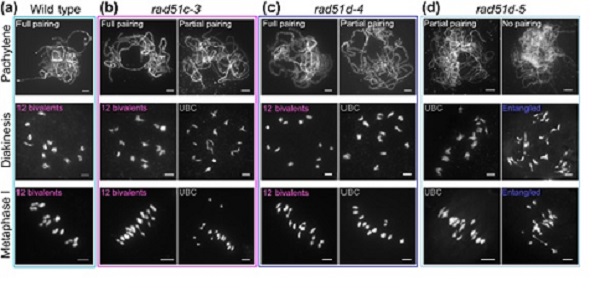
图 1. 三个新突变体的染色体表型
这一研究首次揭示了RAD51旁系同源蛋白在减数分裂重组中的双重功能:在重组早期作为重组酶RAD51和DMC1的招募平台协助DNA单链入侵,在重组晚期与MSH5协同调控重组中间体的解开和交换的成熟 (图 2)。这一功能可能从植物到哺乳动物都是保守的。该研究刷新了研究人员对RAD51旁系同源蛋白的现有认知,并为采用基因编辑手段人工调控重组频率提供了新的基因资源。
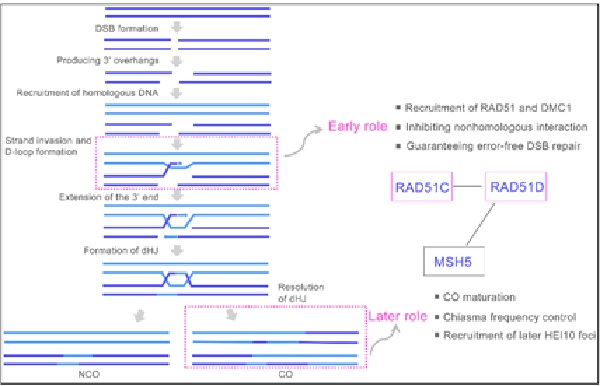
图2. RAD51C和RAD51D在水稻减数分裂中发挥双重作用的工作模式
565net必赢张凡凡博士为该论文的通讯作者及第一作者,山西师范大学石文清博士和中科院遗传发育所博士研究生周月为该论文的共同第一作者,扬州大学程祝宽教授和565net必赢任海云教授对该工作进行了重要指导。该研究得到了国家自然科学基金、中央高校基本科研业务费专项资金以及细胞增殖及调控生物学教育部重点实验室开放课题的资助。
原文链接: https://nph.onlinelibrary.wiley.com/doi/10.1111/nph.19095
